如果我們魔法般地縮小,然後放進到人的大腦中,當她在思考時,我們就會看到所有的「泵」、「活塞」、「齒輪」及「槓桿」工作,我們將能夠完整地描述它們在機械方面的運作,從而完整描述大腦的思想進程。但是,這種描述並沒有提及任何思想!它只不過是對泵、活塞、槓桿進行描述!
——萊布尼茲(1646—1716)
各種領域如何表達自己的原則呢?物理學家使用類似光子、電子、夸克、量子波函數、相關性、節約能源這樣的術語;而天文學家使用恆星、星系、哈勃轉移和黑洞等術語;熱力學家使用熵、第一定律、第二定律、卡諾循環等;生物學家則使用發育、個體發育、DNA和酶。這些術語實際上是每一個專業的代號!一個領域的原則實際上是該領域結構和行為因素交織在一起的集合。
——皮特·丹寧,美國計算機協會前主席,
寫於"Great Principles of Computing"
重要的是,我們要建立處於正確水平的大腦模型。這是當然的,真正科學的模型都是這樣的。雖然理論上化學是基於物理學的,而且可以完全來自物理,但這在實踐中卻難以駕馭,行不通的。因此,化學應該使用它自己的規則和模式。理論上,同樣可以從物理上推出熱力學定律,但這個過程太曲折了。一旦我們擁有足夠數量可以稱為氣體的粒子,而不是說這是一堆粒子,那麼解每個粒子相互作用的方程就將變得不切實際,而熱力學定律此時將變得非常有效。構成氣體的單個分子相互作用是複雜和不可預測的,但由億萬分子組成的氣體本身卻有許多可預見的屬性。
同樣,根植於化學的生物學,也擁有自己的模型。儘管人們在理解一個高水平系統前必須理解水平較低的那個,但往往也沒有必要用低級別的系統動力學的複雜性來表達更高水平的效果。例如,我們可以通過操控動物胎兒的DNA來改變動物的某些特性,而不必瞭解所有DNA的生化機制,更不用說在DNA分子內原子間的相互作用了。
通常情況下,水平較低反而會更複雜。例如一個胰島細胞具有非常複雜的各種生化功能,(其中大部分適用於所有的人類細胞,而有一些適用於所有生物細胞)。然而,在胰島素和消化酶的水平上用數百萬細胞來效仿胰腺的運作過程,雖然並不簡單,但其難度大大小於效仿單一胰島細胞的運作過程。
同樣的問題也適用於大腦的建模和認知,包括從突觸反應的物理現象到神經簇的信息轉換。在那些我們已經成功開發出詳細模型的大腦區域中,我們發現了類似於胰腺中細胞的現象。該模型是複雜的,但仍比單個細胞,甚至是比單個突觸的數學描述還簡單。正如我們前面討論的,這些特定區域建模所需的計算能力大大低於理論上研究所有細胞和突觸的計算能力。
加州理工學院的吉爾斯·勞倫指出:「在大多數情況下,一個系統的集體行為是很難由其組成部分推斷出來的……神經科學是一門系統科學,在這個系統中,需要一階和局部說明圖式,但這還不夠。」隨著我們對每一級的描述和建模的不斷提煉,大腦逆向工程將實現自上而下和自底而上的反覆改良。
受到傳感和掃瞄工具技術未成熟的限制,直到最近,神經系統仍然由過於簡單的模型來描述。這使很多觀察人員懷疑我們的思維過程是否有能力理解自己。皮特·克雷默寫道:「如果想法簡單得足以讓我們明白,那我們就把它想得太簡單了。」50之前,我引用了道格拉斯·霍夫斯塔特將大腦與長頸鹿對比的說法,長頸鹿的大腦結構和人腦沒有多大不同,但顯然它沒有理解自身工作原理的能力。但是,最近在各個層次上設計詳細模型的成功——從像突觸一樣的神經組件到像小腦這樣大的神經區域——都表明建立精確的大腦數學模型,然後用計算模擬這些模型將是一個挑戰,然而一旦數學模型可用,這一任務就會變得可行。雖然神經系統科學建模歷史悠久,但是直到最近,它才變得全面、詳盡,從而能基於此進行模擬大腦的實驗。
子神經型號:突觸和棘
在美國心理學協會2002年的年會上,紐約大學的心理學家和神經學家約瑟夫·勒杜克斯發言說:
如果是記憶塑造了真實的我們,並且記憶是大腦的一項功能,那麼突觸(神經元通過它相互溝通,同時它也是記憶編碼的物理結構)就是最基本的單位……突觸在大腦組織過程中的地位非常低,但我認為它們非常重要……它們自身的總和是大腦子系統,其中每一個都有自己的記憶形式以及子系統之間複雜的相互作用。如果沒有突觸可塑性(突觸連接強度可調節的特性),這些系統學習所必需的改變就將不可能發生。51
雖然早期的模型認為神經元是傳輸信息的主要單位,但是現在已經轉向強調其亞細胞成分。例如,計算神經科學家安東尼·貝爾認為:
分子和生物物理過程控制引入峰電位的神經敏感性(包括突觸的效率和突觸後響應性),產生峰電位的神經元興奮性,以及它產生的峰電位模式和新突觸形成可能性(動態重新布線),使之只列出四種最明顯的亞神經一級的干擾。此外,類似本地電場和一氧化氮跨膜擴散這樣的跨神經元活動也有影響,還有相關的神經元放電和為細胞供給能量(血流量),而後者直接與神經活動有關。這個清單還可以繼續。我相信,任何人認真研究神經調節、離子通道或突觸機制,都不能將神經元的水平作為一個單獨的計算水平,哪怕發現它是一個有用的描述水平。52
事實上,真實的大腦突觸比邁克洛-匹茲所描繪的經典神經網絡模型要複雜得多。突觸的反應受到一系列因素的影響,包括多種離子勢能(電壓)和神經遞質所控制的多通道活動和神經調節。雖然過去的20年已經有了巨大的發展,但是,在發展中,數學公式總是優先於神經元,樹突和突觸的行為以及脈衝系列的信息表現形式(脈衝被神經元激活)。最近,皮特·達揚和拉裡·阿爾伯特編寫現有的非線性微分方程的概要,其中描述了通過成千上萬的實驗所得到的知識。53關於神經元體和突觸的生物物理學,以及神經元正反饋網絡的行為,這些都存在著經得起論證的模型,就像在視網膜、視神經以及其他各種神經元中發現的那樣。
對神經突觸工作機制的關注起源於海勃的開創性工作。海勃提出了這個問題:短期(也稱為「正活動的」)記憶是如何工作的?與短期記憶相關聯的大腦區域是前額葉皮層,雖然我們已經意識到,短期信息保留的不同形式已經在大多數其他已被仔細研究過的神經回路中得到證實。
海勃的大部分工作集中在改變突觸的狀態(以此來加強或者抑制對於信號的接收)和更有爭議的反射神經回路(其中包括在連續回路內的神經放電)上。54海勃提出的另一個理論是,神經元細胞自身進行狀態改變,也就是說,記憶功能在細胞軀體(身體)內。實驗證據支持了這些模型的可能性。經典海勃突觸記憶和反射記憶在記錄信息可用之前需要一段時間延遲。實驗表明,至少在大腦的某些區域,神經反應太快以致不能使用標準的學習模式,只能通過誘髮型學習來完成。55
海勃沒有直接預想到的另一種可能性是神經元自身連接的實時變化。最近的掃瞄結果表明了樹突峰電位和新突觸可以迅速增長,因此必須重視這一機制。實驗還表明,在突觸水平上的大量豐富的學習超越了簡單海勃模型。突觸可以很快改變其狀態,但由於受到不斷刺激,或缺少刺激,又或是其他原因,它們又開始緩慢地衰減。56
雖然現代模式比海勃設計的簡單模型要複雜得多,但他的直覺大部分已經被證明是正確的。除海勃的突觸可塑性外,目前的模型還包括提供調節功能的全局進程。例如,通過突觸縮放使突觸勢能避免變成零(否則就無法通過乘法的方法增加)或變得過高,從而能主導網絡。在體外實驗中,我們已經在上文所描述的神經網絡、海馬體及脊髓神經中發現了突觸縮放,57其他機制對於整體峰電位週期和突觸勢能的分佈是很敏感的。仿真證實了最近發現的機制具有提高學習和穩定網絡的能力。
在認識突觸過程中,最令人激動的新發展是,突觸的拓撲和它們之間的連接在不斷變化。通過一個新穎的掃瞄系統,我們第一次看到了突觸連接的迅速變化。這個系統需要一個轉基因動物,其神經元可以發出螢光綠。該系統可以刻畫活的神經組織,並且具有足夠高的分辨率以捕獲樹突(中間的連接)以及棘:萌發自樹突的微小突起,它是突觸的初級形式。
神經生物學家卡瑞爾·斯沃博達和他在長島冷泉港實驗室的同事將該掃瞄系統用於老鼠,從分析鬍鬚信息來調查分析神經網絡,這項研究描述了神經學習的迷人景象。樹突在不斷地長出新棘。其中大多數只持續了一兩天,但有時棘也會保持穩定。「我們相信看到的高周轉量會在神經可塑性上發揮重要作用,其中萌發的棘伸向臨近的神經元以探查不同的突觸前搭檔,」斯沃博達如是說,「如果給定的連接是有利的,也就是能反映一種可取的大腦重新布線,那麼這些突觸將是穩定的並且會更加永久化。但這些突觸大多不是朝著正確的方向,而且它們是可伸縮的。」58
另一個已經觀察到的一致現象是,如果某一特定的刺激重複發生,神經反應會隨時間而減少。這種適應性給予新的刺激模式最高的優先級。紐約大學醫學院的神經生物學家甘文彪做了相似的工作,他對成年小鼠視覺皮層神經棘的研究表明,這種棘機制可以容納長期記憶。「假設10歲的孩子使用1000條連接存儲一條信息。當他80歲時,不管情況如何變化,這些連接的1/4將依然存在。這就是為什麼你還記得你童年的經歷。甘還解釋說,」我們的想法是,當你學習或記憶時,你實際上並不需要許多新的突觸,也不需要去除舊的那些。你只需要修改先前存在的突觸的強度來進行短期的學習和記憶。但是,很可能是產生或消除一些突觸,以實現長期記憶。59
之所以記憶可以保持完整(即使3/4的連接已經消失),是因為該編碼方法的特性類似於全息圖。在全息圖中,存儲的信息廣泛地散佈在整個區域內。如果你銷毀了全息圖的3/4,儘管只剩1/4,但是整個圖像仍將保持不變。紅木神經科學研究所的神經科學家彭蒂·卡內爾瓦的研究支持了這一觀點:記憶動態地分佈在神經元的整個區域。這就解釋了為什麼老年人的記憶可以延續下去,只是表現得越來越模糊,因為它們的分辨率已經削弱。
神經模式
研究人員還發現,特定的神經元執行專門的識別任務。雞的腦幹實驗檢測到聲音到達而兩耳時會有一段特殊的延遲。60不同神經元的時間延遲也會不同。儘管這些神經元工作時總有許多複雜的非常規行為,但它們實際完成的內容還是很容易描述的,並且也很容易複製。美國加州大學聖地亞哥神經學家斯科特·馬克格如是說:「最近的神經生物學實驗的結果顯示了精確同步在學習和記憶中的重要作用。」61
電子神經元。加利福尼亞大學在聖地亞哥的研究所最近進行了一項關於非線性科學的實驗,該實驗揭示了模仿生物神經元所需的電子勢能。神經元(生物或其他的)是混沌計算的最好例證。每個神經元的行為在本質上都是不可預知的。當整個神經元網絡接收輸入(來自外面的世界或是其他神經細胞網絡)時,一上來信號似乎都很狂亂而且很隨機。隨著時間推移,通常是幾分之一秒左右,神經元間混沌的相互作用慢慢消逝,神經放電開始呈現出穩定的格局。這一模式顯示了神經網絡的「決策」。如果神經網絡正在執行模式識別任務(這些任務構成了人類大腦的活動),那麼這一突發模式就代表了正確的識別結果。
因此,聖地亞哥的研究人員提出的問題是:電子神經元能否在生物神經元旁邊與之攜手共舞?他們把人工神經元和生物神經元放在一個網絡中,這一生物和非生物混合的網絡按同樣的方式執行(混亂的相互作用之後進入一個穩定的突發模式),而其結果和全生物網絡是一樣的。從本質上說,生物神經元接受了它們的電子同行。這表明這些神經元的混沌數學模型相當準確。
大腦可塑性
1861年法國神經外科醫生保羅·布洛卡將大腦的受傷或外科手術影響的大腦區域與某一失去的技能聯繫起來,如精細的動作技能或語言能力。一個多世紀以來,科學家認為這些區域是完成具體任務的硬件。雖然某些腦區域往往用於特定類型的技能,但我們現在明白,遇到如中風一樣的傷害時,大腦分配的任務可以做出一些轉變。在1965年的經典研究中,胡貝爾和維塞爾展示了在神經系統受到如中風一樣的損壞時,大腦廣泛而深遠的識別功能仍然可以進行。62
此外,在某一給定區域,連接和突觸的詳細佈置是該區域使用程度的直接產物。由於大腦掃瞄已達到足夠高的分辨率,已經足以檢測樹突棘的增長和新突觸的形成,我們可以看到大腦生長並一步步地看著想法的產生(見圖4-3)。這使笛卡爾的名言「我思故我在」的意義又有了新的色彩。
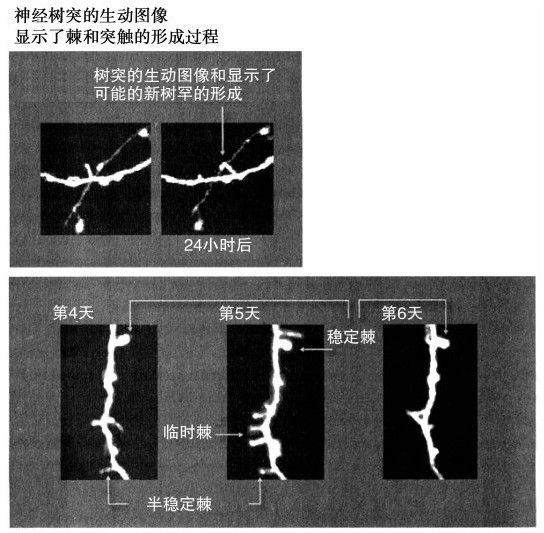
圖 4-3
在舊金山的加利福尼亞大學,邁克爾·莫山尼奇和他的同事進行了一項猴子的實驗,他們將食物放置在一個猴子必須巧妙活動一個手指才能獲得的位置。對這一活動之前和之後的腦部掃瞄顯示,在大腦中負責控制該活動的區域中,連接和突觸急劇增長。
阿拉巴馬大學的愛德華·塔布研究了腦皮層區域對手指觸覺輸入的反應。比較不太懂音樂的人和老練的樂器演奏者,可以發現專門控制右手手指的大腦區域並沒有什麼差異,但控制左手手指的區域卻有著很大的不同。
如果繪製出基於觸摸分析的腦組織量的圖像,我們會發現控制音樂家左手的腦組織量非常大。儘管與從小就練習絃樂器的音樂家相比,差異會更大,但「即使你是從40歲才開始練習小提琴,」塔布說,「你仍然可以獲得腦重組。」63
類似的發現來自對一款軟件程序的評價,該軟件由羅格斯大學的保羅·塔萊和史蒂夫·米勒製作,稱為Fast ForWord(快速朗讀),旨在幫助有閱讀障礙的學生。該程序為兒童朗讀文章,並減緩斷音的發聲,如"b"和"P",這是由於很多誦讀困難的學生在聽到快速發音時,很難察覺這些聲音。而通過這種形式的修改,這一軟件很好地幫助了這些兒童對朗讀的學習。斯坦福大學的約翰·加布裡埃利發現,利用fMRI,與語言處理有關的大腦左側前額葉區域確實有所增長,尤其是這些誦讀困難的學生,在使用該軟件時增長更多。塔萊說:「你通過獲得的輸入來創造你的大腦。」
甚至不需要通過物理活動表達一個人的思想就可以驅使大腦重新布線。哈佛大學的阿爾瓦羅·帕斯誇爾-利昂博士對進行簡單鋼琴練習前後的志願者的大腦進行了掃瞄,志願者大腦運動皮質的變化是他們練習的直接結果。然後他開始了另一組實驗,在這一組中,他讓志願者僅僅想像一些鋼琴的練習而沒有任何實際的肌肉運動。而該組志願者的運動皮層卻產生了與第一組相同的顯著變化。64
最近關於視覺空間關係的fMRI研究發現,神經元之間的連接在一個單一的學習會話期間會迅速變化。研究人員發現,這種變化發生在後頂葉皮層細胞(在所謂的「背」通路中,包含位置和視覺刺激的空間屬性的信息)和後顳葉皮層(在「腹」通路中,其中包含了不同層次抽像的認知不變特徵)。65很明顯,那種變化率與學習速率成正比。66
加州大學聖地亞哥分校的研究人員給出了關於短期記憶和長期記憶形成差異的重要觀點。利用高分辨率掃瞄手段,科學家可以清楚地看到海馬體(與形式長期記憶有關的大腦區域)的神經突觸細胞的化學變化。67他們發現,細胞第一次受到刺激時,肌動蛋白(一種影響神經系統的化學物質)會向與神經突觸連接的細胞移動。這也刺激了相鄰細胞的肌動蛋白遠離被激活的細胞。這些變化僅僅會持續幾分鐘,然而,如果得到了充分的反覆刺激,那就會有更顯著和更持久的變化發生。
「短期變化僅僅是神經細胞之間相互聯繫的正常方式中的一種。」主要作者邁克爾·克裡斯克說:
要使神經元產生長期變化,必須在這一小時內四次刺激神經元。神經突觸會分裂並且會有新的神經突觸形成,進而產生一個長期的變化,並可能持續一生。類比於人的記憶,當你看到或聽到一件事,它可能會停留在腦海中幾分鐘。而如果它不重要,就會逐漸在我們腦海中消失,10分鐘後我們就會忘記它。但如果我們再次聽到或看到它,並且這件事在接下來的一個小時內一直持續發生,你可能就會記住它更久一些。而事情如果發生了很多次,你就會一生都記住。一旦你拿一個軸突形成兩個新的連接,這些連接就將會非常穩定,沒有理由相信它們會消失。這就是那種將會持續一生的改變。
「這就像一堂鋼琴課」,作者之一的生物學教授夕紀子·高達說,「如果你將一首音樂彈了一遍又一遍,它將扎根於你的記憶中。」同樣,神經科學家威爾和辛格發表在《科學》雜誌的一篇文章說到,在視覺皮層中他們找到了新的神經元連接動態生成的證據,就像唐納德·海勃描述的"What fires together wires together"。68
另外一個關於記憶形成的觀點來自Cell(細胞學)上的一篇研究報告。研究人員發現,CPEB69蛋白質實際上是通過改變它們的突觸形狀來記錄存儲信息的。最讓人吃驚的是,當CPEB處於朊粒態時才能執行存儲功能。
「我們暫時粗略地瞭解了記憶的工作原理,但是對於這種存儲設備,我們至今沒有一個清新的概念。」合著者、懷特海德生物工程研究院的負責人蘇珊·林德基斯特說,「這項研究揭示了這一存儲設備可能的存在形式,但令人驚訝的是,我們發現這一過程還涉及朊粒的活動。這說明朊粒不僅具有奇異的特點,而且可能參與生物體基本的處理中。」就像我在第3章講到的,人類工程師研究發現朊粒具有產生電子存儲功能的強大能力。
腦掃瞄研究也揭示了抑制不需要的和不良的記憶的機制,這一發現使西格蒙德·弗洛伊德大為高興。70利用fMRI,斯坦福大學的科學家要求被研究人員試圖去忘記他們早期已經記憶的信息。在此活動期間,與抑制記憶相關的額葉皮層區域表現得相當活躍,而海馬體這一通常與記憶相關的區域卻相對不太活躍。這些研究結果「確定了一個積極遺忘進程的存在,並為引導動機性遺忘的研究建立了一個神經生物學模型,」斯坦福大學心理學教授約翰·加布裡埃利和他的同事這樣寫道。加布裡埃利還評論說:「最大的新聞是,我們已經證實了人類的大腦如何阻止不需要的記憶,存在這樣一種機制,並且它具有生物學基礎。它使你不再去考慮這樣的可能性,即大腦裡不存在任何事物可以抑制記憶——這其實一直是被曲解的謬論。」
除了使神經元之間產生新的連接,大腦也可以通過神經干細胞產生新的神經元,其中神經干細胞通過複製來保持自身的供應。在複製的過程中,一些神經干細胞變成了「神經前體」細胞,它們輪流發展成為兩類被稱作星形膠質細胞和少突膠質細胞的支持細胞。這些細胞進一步演變成為特定類型的神經元。
然而,這些分化不會發生,除非神經干細胞從它們在腦室中的原始位置離開。只有大約一半的神經細胞成功完成該過程,這類似於妊娠和早期幼兒階段,大腦只有一部分生長中的神經元存活下來。科學家們希望能夠通過直接向目標區域注射神經干細胞從而繞過神經遷移過程,或者創造藥物以促進神經形成的進程(產生新神經細胞)來修復由於疾病或損傷而對大腦造成的傷害。71
一項由美國索爾科生物學研究所的遺傳學研究人員弗雷德·戛格、坎貝爾曼和亨利埃特·范·普拉赫進行的實驗表明,神經形成實際上是受我們的生活經歷刺激的。將小鼠從無菌、乏味的籠子裡移動到一個有刺激性的籠子裡,它們海馬區的分裂細胞大約會增加一倍。72
大腦區域建模
人類大腦最可能是由大量相對較小的分佈式系統組成,這些系統按照胚胎學排列成複雜的集合,並由之後增加的串行符號系統進行部分(也只是部分)控制。但是那些做了底層大部分必需工作的子符號系統,由於其本身的性質,阻礙了大腦的所有其他部分理解它們的工作原理。這本身就可以幫助我們解釋,為什麼人們做了如此多的事情,卻仍然只掌握了一些關於如何完成這些事的不完整想法。
——馬文·明斯基和西摩·帕勃73
常識不是一件簡單的事。相反,它是一個包含著來之不易的實踐經驗的龐大集合體——包括眾多生活教訓、異常事件、處理方式、個人傾向、還有平衡和檢驗。
——馬文·明斯基
除了關於每個大腦區域組織可塑性的新見解,研究人員還在迅速創建大腦特定區域的詳細模型。這些神經形態模型和模擬仿真只是稍稍滯後於它們所基於的信息可用性。由對神經掃瞄得到的神經元和神經元間的連接數據的研究到創建有效模型和可行的模擬仿真,這一轉變的快速成功淹沒了經常提及的關於理解我們自己大腦內在能力的懷疑論。
對基於非線性對非線性和突觸對突觸的人腦功能進行建模並不是必要的。在單個神經元和連接(例如,小腦)中,對存儲記憶和技能的區域模擬確實要使用詳細的細胞模型。然而,即使在這些區域,模擬所需的計算量也比所有神經元組件所暗含的要小得多。我們將在接下來描述的細胞模擬中看到這一真實性。
雖然在大腦的上億個連接中,每個神經元的亞神經元部分都有著大量的詳細的複雜性和非線性,以及混亂的半隨機布線圖,但在過去20年中,在數學建模方面還是取得了重大進展,例如自適應非線性系統。通常沒有必要去保存樹突的枝的確切形狀和每個神經元連接的精確「波浪線」。我們可以通過對其進行適當水平上的動態分析來理解大腦廣闊區域的運作原則。
我們在大腦區域的建模和模擬上已經取得了重大成功。對這些模擬進行測試,並將得到的數據與對真實人腦進行心理物理學實驗所獲得的數據進行比較,由此取得了令人矚目的成果。考慮到我們當前相對粗糙的掃瞄檢測技術和傳感工具,在建模上取得的成功(如以下正在進行的工作),證實了從大量採集來的數據中提取正確觀點的能力。接下來介紹的是大腦區域成功建模中的幾個例子,所有工作都還在進展中。
神經形態模型:小腦
我在《The Age of Spiritual Machines》中發現了這樣一個問題:一個10歲小孩怎樣成功抓住一個高飛球?74一個孩子能看到的全部信息只是球的位置軌跡。要真正推斷球在三維空間的路徑,需要解複雜的微分方程組,還需要解附加的方程來預測球的未來方向,以及解更多的方程來將結果轉化為球員自己的運動。一個這麼小的外場手是如何在沒有電腦幫助,也沒有練習過解微分方程的情況下,在如此短的時間內完成所有這些呢?顯然,他並沒有意識地去解方程,但是他的大腦是如何解決這一問題的呢?
自從《The Age of Spiritual Machines》出版以來,我們對這一技能形成的基本過程的理解有了很大的發展。正如我所假設的,這個問題並不是通過建立三維運動的心理模型來解決。相反,問題可以這樣解決,直接將觀察到的球的移動翻譯成運動者的適當移動,並改變其胳膊和腿的位置。羅切斯特大學的亞歷山大·博基特和華盛頓大學的勞倫斯·斯奈德描述了數學的「基礎功能」能代表這種直接轉化,即直接將視野中感知到的運動轉化為所需的肌肉運動。75此外,對於最近取得進展的小腦功能模型的分析,證明了我們小腦的神經回路確實具有學習能力,然後通過運用基礎功能實現這些感覺運動的轉換。當我們採用逐步逼近的學習過程去執行一個感覺運動的任務時,如抓住高飛球,我們就是在訓練小腦的突觸學習適當的基礎功能。小腦通過這些基礎功能可以執行兩種類型的轉換:判斷一個理想的結果是否可行(為「逆向內部模型」)和從可能的動作集中預測一個結果(「前進內部模式」)。托馬斯·波吉奧指出,基本功能的概念描述了大腦的學習過程,這一點超越了電機控制。76
大腦區域有一塊區域,其顏色由灰色和白色組成,有棒球大小,豆型,這一區域被命名為小腦,位於腦幹之上,它包括超過大腦一半以上的神經元。小腦提供了很多方面的關鍵功能,包括感覺運動協調、平衡,控制運動的功能,並有能力預測行動的結果(我們自己以及其他動物和人)。77儘管其功能多種多樣,但是它的突觸和細胞組織極其一致,只涉及幾種類型的神經元細胞,看上去它似乎只是在完成一種特定類型的計算。78
儘管小腦的信息處理方式是同樣的,但是其廣泛的職能可以通過從大腦皮層(通過腦幹細胞核,然後通過小腦的苔狀纖維細胞)和其他區域(特別是大腦的「下橄欖核」區域,通過小腦的攀緣纖維細胞)接收的各種輸入來進行理解。小腦負責理解我們感覺輸入的時間和順序,同時控制我們身體的運動。
小腦也是說明大腦如此大的容量是怎樣大大超過了它的緊湊基因組的一個例子(見圖4-4)。大多數基因組致力於大腦對於各種神經細胞(包括樹突、棘和突觸)的詳細結構的描述以及這些結構如何應對刺激和變化。相對的,幾乎沒有基因代碼負責實際的「布線」。在小腦中,基本的布線方法被重複數十億次。顯然,沒有基因組提供有關每個小腦結構每一次重複的具體信息,相反,它們提供了具體的某些約束,對這些結構怎樣重複(正如基因組沒有指定其他器官細胞的確切位置一樣)制定了一些限制。
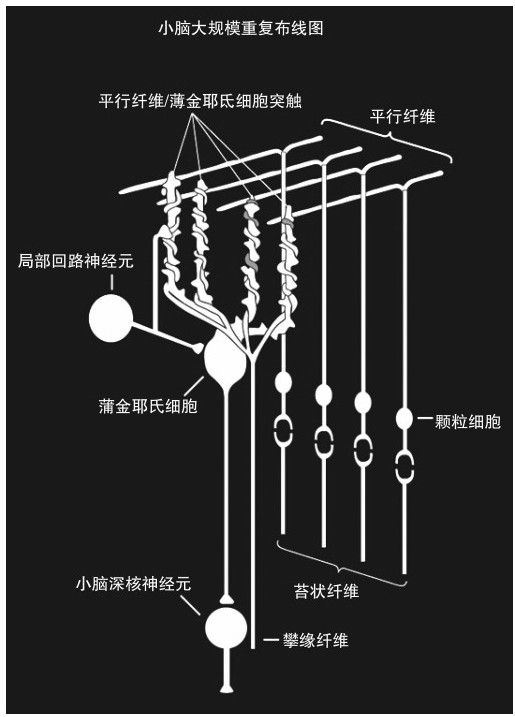
圖 4-4
小腦的產出輸送到大約20萬α運動神經元中,從而確定最終的信號來控制身體中大約600塊肌肉組織。輸入到α運動神經元的物質不會直接指定每個肌肉組織運動,而是進行更緊湊地編製,因此很難明確理解這一過程。指定肌肉運動的最終信號由更低級的神經系統決定,特別是腦幹和脊髓。79有趣的是,這個組織在章魚的身體裡使用到了極限,中樞神經系統向它的每個觸角發出高層指令(如「抓住這個物體,把它拿得近一些」),將信號傳遞給每個觸角內的外圍神經系統來執行該任務。80
最近幾年,我們對小腦的三種主要神經類型有了很多認識。稱作「攀緣纖維」的神經元提供信號來訓練小腦。小腦的大多數輸出結果都來自蒲金耶氏細胞(是以約翰內斯·蒲金耶氏的名字命名的,他於1837年發現了這種細胞),每一個蒲金耶氏細胞可以接收大約20萬的輸入(突觸),而普通的神經元一般只能接收10萬左右。大部分的輸入都來自於顆粒細胞,這種細胞是最小的神經元,大約每平方毫米600萬個(見圖4-5)。在研究小孩學習寫字時小腦作用的試驗中,研究人員發現蒲金耶氏細胞決定著移動的序列,每一個細胞影響一個具體的移動動作。81很明顯,小腦需要從視覺皮質不斷獲得視覺感知。研究人員將小腦細胞的結構和實驗觀察關聯起來發現,在寫字時,移動軌跡的曲率和移動的速度成反比。就是說,當你寫字時,畫直線的移動速度明顯比畫曲線時快得多。
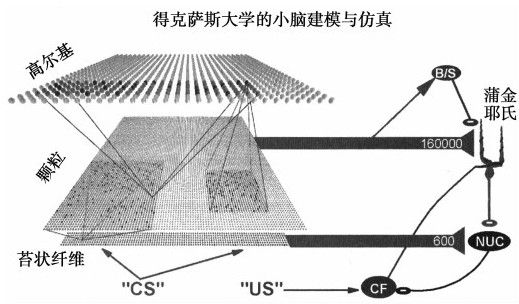
圖 4-5
複雜細胞研究和動物研究為我們提供了關於生理學和小腦突觸組織,82輸入和輸出的數據解碼,以及執行轉變的詳細描述。基於多項研究得到的數據,哈維爾·麥迪納、邁克爾·馬克和他在得克薩斯州醫學院的同事們為小腦設計了一種自底向上的詳細模擬。83它具有一萬個模擬神經元和三萬個神經突觸,並且它包括了小腦中所有重要類型的細胞。84這些細胞和神經突觸的連接由一台計算機控制,這台電腦依據一些限制和規則進行「布線」,從而模擬小腦區域的功能。這一點與遺傳序列控制真實人腦布線的隨機(有限制條件的隨機)方法類似。85而想把得州大學的小腦模擬擴展成一個更大規模的模型也並不困難。
得州大學的研究者們為他們的神經模擬提供了重要的研究經驗,並將實驗結果與很多關於真實人類的相似實驗進行了對比。在人類研究中,我們進行了這樣的實驗:在發出音響的同時對被研究人員的眼瞼吹一陣空氣,這樣人的眼瞼就會關閉。如果結合空氣和音響這兩種因素進行100~200次實驗,這名參與者就會學到這種關聯,而之後僅僅是聽到音響,就會閉上眼睛。而如果之後又採用只有聲響而不再對眼睛吹氣,那麼該參與者將最終分離這兩種刺激(並會「結束」這種響應),所以學習是雙向的。之後經過調整各種參數,使模擬能夠合理匹配人和動物小腦調節的實驗結果。有趣的是,研究人員發現,如果他們模擬小腦損傷(刪除部分模擬小腦網絡),結果和使兔子小腦真實損傷實驗得到的結果也很相似。86
由於這一大塊大腦區域的一致性以及與之相關的神經元間布線的簡單性,其輸入輸出的轉變與其他區域相比要容易理解一些,儘管這些關聯方程式還需要繼續精煉改進,但這一自下而上的模擬確實令人印象深刻。
另一個例子:瓦特的聽覺區域模型
我相信創造一個類人腦智能的方法就是建造一個實時工作模擬系統,足夠詳細和準確地表達每個需要執行的計算的本質,並能對該實時系統進行度量以核實其操作的正確性。之所以要求實時系統是因為我們不得不去處理那些不方便的、複雜的真實世界的輸入,而這些輸入我們以前並沒有考慮如何去表達。這一模型一定要在足夠的分辨率下運行,這樣才能與實時系統相匹配,所以我們對於每一階段所代表的信息都建立了正確的直覺知識。根據密德的說法,87這一模型有必要從實時系統中已經理解得很好的系統邊緣(例如傳感器)開始,之後進入理解得少一些的區域……通過這種方法,這一模型可以從根本上促進我們對這一系統的理解,而不是簡單地反映已具備的理解水平。在這種極其複雜的情況下,也許唯一能夠理解實時系統的實際方式就是建立一個工作模型,隨著不斷發展,在傳感器內部建立我們新的有效能力——形象化系統的複雜性。這種做法可以稱作大腦逆向工程……請注意,我不是在倡導盲目地複製這些我們不明白其目的的結構,就像故事中的那樣,伊卡洛斯天真地建造了羽毛和翅膀,以為這樣就可以飛翔。相反,我主張尊重已經是在較低水平被很好理解和認識的複雜性和豐富性,只有這樣我們才能向更高的水平進發。
——勞埃德·瓦特88
關於大腦區域神經形態建模的一個重要例子就是勞埃德·瓦特和他的同事們開發的人類聽覺處理系統的一個關鍵部分的綜合複製(見圖4-6)。89它基於特定神經類型的神經生物學研究以及神經元連接信息。該模型與人類聽覺有著許多相同的屬性,可以進行聲音來源定位和聲音識別,它有5條聽覺信息處理的平行通路,包括神經處理每一階段信息的中間表示法。瓦特按照實時計算機的操作實驗他的模型,雖然這項工作仍在進行當中,但它已經闡明了神經生物學模型和大腦連接數據轉化為仿真的可行性。這一軟件不是像已經描述過的小腦模型那樣基於複製單個神經元和連接,它是基於每一區域所執行的轉換。
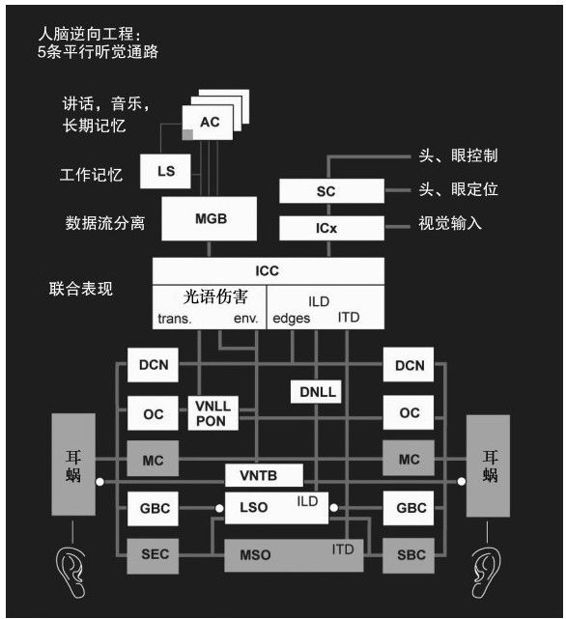
圖 4-6
瓦特的軟件已經能夠匹配人類的聽覺和辨音能力小實驗中所顯露出的錯綜複雜的關係。瓦特使用這一模型作為語音識別系統的預處理(前端),已經證實可以從背景音中識別出一位發言者(「雞尾酒會效應」)。這是一個了不起的壯舉,這將實現人類有能力做到而在自動語音識別系統中卻一直未實現的功能。90
就像人的聽力,瓦特的耳蝸模型被賦予了光譜靈敏度(我們在某個固定的頻率範圍能夠聽得更好)、時間回應(我們對於聲音的傳送時間很敏感,它使我們感知到聲音來源的空間位置)、掩蔽、非線性頻率相關的振幅壓縮(允許更大的動態範圍,即同時聽到響亮和安靜的聲音)、增益控制(擴增),以及其他微妙的特徵。它獲得的成果可以直接由生物和心理物理數據給出驗證。
該模型的下一個部分是耳蝸神經核,耶魯大學神經科學和神經生物學教授戈登·謝福德91將其描述為「大腦內理解得最好的區域之一」。92瓦特的人工耳蝸神經核仿真模型正是基於楊戈的工作,楊戈詳細描述了「這些並不可少的細胞類型,旨在探測頻譜能量,寬帶瞬變和光譜波道的微調,提高暫時包圍在光譜波道、光譜邊緣和缺口內的靈敏度,以及所有在尖峰神經編碼的有限動態範圍內再次調整而獲取的最優靈敏度」。93
瓦特模型還捕捉到了許多其他的細節,例如由中間上橄欖體細胞計算出來的兩耳時間差(lTD)94。除此之外,它還能描繪由側面上橄欖體細胞計算出的雙耳位准差(ILD),以及下丘細胞產生的規範化和調整方法。95
視覺系統
我們在視覺信息編碼上已經取得了足夠的進步,實驗性的視網膜移植已經取得了很大進展並可以利用外科手術移植到病人的眼部。97但是由於視覺系統的相對複雜性,我們對於視覺系統的認識還遠遠不如對於聽覺區域的認識深刻。我們有初步的兩個視覺區域(分別稱為VI和MT)執行轉換的模型,儘管它們不在視覺神經層。另外還有其他的36個視覺區域,我們需要在很高的解析度下或者是利用精確的傳感器才能夠掃瞄這些更深層的區域,以便查明它們各自的功能。
麻省理工學院的托馬斯·博吉奧是視覺處理認知的先驅,他將自己的工作分為兩種:識別和分類。98根據他的解釋,前者相對來說比較容易理解,我們已經設計出能夠成功識別人臉的實驗版和商業版系統。99這些系統已經成為安全系統的一部分,用來控制職員的出入的門禁,另外,在提款機中也有應用。分類,即辨別如人和汽車之間或狗和貓之間的不同之處,這是一個比這些更複雜的工作,儘管現在已經取得了一些進步,但還是遠遠不夠的。100
視覺識別系統的早期階段(根據進化論)很大程度上是一種前饋系統(沒有反饋),這個系統能夠檢測到日益複雜的事物特徵。博吉奧和馬克西米蘭·瑞斯胡博爾寫道:「獼猴的後顳葉下皮質的一個單一神經元可能會變成……具有成千上萬複雜形狀的字典。」有證據表明視覺識別在識別過程中(包括MEG研究)使用了前饋系統,並顯示人類視覺系統檢測物體時需要大約150毫秒。這和顳葉下皮質的功能檢測細胞的潛在因素相匹配,所以在早期的決策階段,反饋作用的時間看上去並沒有發揮很大的作用。
最近的實驗已經開始採用分等級的方法,由此檢測的特徵就可以在系統的後期階段進行分析。101從對獼猴的研究得知,經過訓練的獼猴,其顳葉下皮質的神經元好像對物體的複雜特徵會做出回應。儘管大部分的神經細胞只是對所看到的物體的特殊景像有反應,還有一些則是不管看到什麼景象都有反應。其他關於獼猴視覺系統的調查還包括研究一些細胞的特殊類型、連通性模式,以及對於信息流的高水平描述。102
很多的文獻資料表明,在很多更複雜的模式識別任務中,我稱為「假設和檢驗」的方法是很有效的。例如,首先猜測觀察到的事物究竟是個什麼東西,然後再檢驗你的假設和你的實際觀察有多大的匹配度。103我們經常更關心的是假設,而不是事後的檢測,這也就解釋了為什麼人們常常寧願相信他們假設看見和聽見的事物而不是實際存在的事物。當然,「假設和檢驗」在我們以計算機為基礎的模式識別系統中也非常有用。
儘管我們常常錯誤地認為可以眼睛中獲得高分辨率的圖像,而實際上,視神經傳遞給我們的只是視覺區域感興趣點的大致輪廓和蛛絲馬跡。然後我們通過皮質記憶(用來解釋一系列平行通道上極端低分辨率的影像)來製造這個世界的幻象。在2001年的《自然》雜誌發表的一篇文章上,加州大學伯克利分校的分子與細胞生物學教授弗蘭克·維布林和他的博士生波頓·羅斯卡,M.D.向我們展示了視神經所承載的10~12個輸出通道,它們每一個都只承擔已給出景象的很少信息。 104我們稱為神經節細胞的其中一組只是傳遞了一些關於物體邊緣輪廓的信息(changes in contrast),另一組則具有檢測大面積相同顏色的功能,而第三組只是對於興趣點背後的背景比較敏感(見圖4-7)。
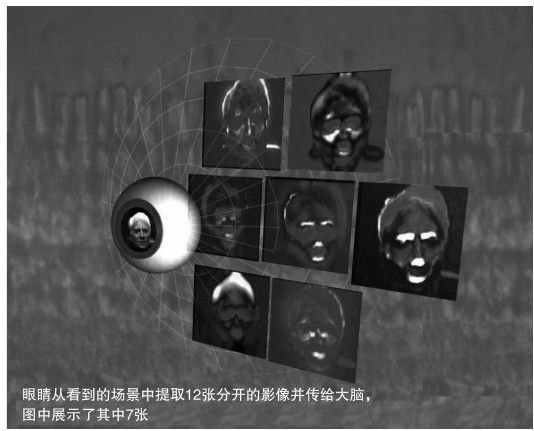
圖 4-7
維布林說:「儘管我們認為我們如此完整地看到了這個世界,而事實上,我們所看到的也僅僅只是一些暗示性的信息以及時間和空間的輪廓而已。這12張圖像是我們一直以來所獲得的周圍事物的所有信息,它們是如此稀疏,而我們正是利用這些重建了我們豐富多彩的視覺世界。我很好奇,為什麼自然最終會選擇這麼簡單的十二種影像,而它又是如何向我們提供我們所需要的這個世界的全部信息的呢?」這樣的發現對於智能系統的發展肯定具有很大的推進作用,而在這種系統中,眼睛以及早期的視覺處理將會被取代。
在第3章中,我提到的了機器人技術的先行者漢斯·莫拉維茨,他已經通過視網膜神經和大腦中早期的視覺處理轉變了工程學上的圖像處理。漢斯·莫拉維茨用了30年的時間致力於建造一個系統,期望這個系統能夠趕上人類視覺系統的能力,從而達到從視覺上重建世界的目標。而它只是在最近才有足夠的處理能力,因為微處理器已經能複製人類級別的特徵檢測,而莫拉維茨正在應用他的計算機模擬新一代的機器人,使之能夠通過人類水平的視覺能力在複雜的環境中自動導航。105
卡福·密德一直引領著特殊神經芯片的研究,它將晶體管使用於它們天然的模擬模式,這一模式可以提供非常有效的神經處理的模擬仿真。密德展示過這樣一個芯片,此芯片可以應用該方法模擬視網膜的功能,並進行視覺神經早期轉換。106
檢測移動是視覺識別的一種特殊類型,它是杜平根生物物理研究所的重點研究領域。最基本的研究模型很簡單:將一個感受器上的信號和在其鄰近感受器的延遲信號相對比。107這一模型要求速度在特定範圍內,如果超過某一特定速度,則會導致令人驚訝的結果。這時,當觀察對象的速度繼續增加,移動探測器響應靈敏度將減少。動物實驗結果(基於行為和我的分析,以及神經產出)和人的實驗結果(基於報道的看法)都與該模型很匹配。
正在行進中的其他工作:人造海馬體和人造小腦
海馬體對於新信息的學習和長期的記憶存儲都具有很關鍵的作用。南加州大學的特德·伯傑和他的同事們通過數萬次的電信號刺激老鼠的海馬體切片,從而繪製出該區域的信號模式,以此來確定哪些輸入可以產生相對應的輸出。108隨後他們開發出了一種模擬這種結構的實時數學模型,這種數學模型模擬了海馬體的信息轉換,並且他們已經把這個程序固化到一個芯片上。109為了測試這一芯片,他們計劃首先破壞動物原有的海馬體,並記錄產生的失敗存儲,然後將海馬體芯片安裝到被破壞的區域,並檢測動物的記憶能否在芯片上重現。
最終這種方法能夠用來取代患傷風、癲癇或者阿爾茲海默症的病人的海馬體。這種芯片將被植入病人的頭蓋骨上,而不是大腦內,它能夠通過兩列導線和大腦進行通信,以取代損壞的海馬體區域的功能。其中一列導線是用來接收大腦傳來的電信號,另一部分則是用來向大腦回饋相應的信息。
另一個被模式化和被模擬的是人類的小腦區域,它負責調節人的平衡和協調四肢的運動。該國際研究小組努力的目標是將他們的人工小腦電路應用到軍用機器人以及一般的機器人中,以便有效地協助殘疾人。110選擇這一特定的大腦區域的一個原因是:「這是目前在所有脊椎動物都具有的部分,不管是在最簡單還是最複雜的大腦中,它都是大同小異的,」魯道夫·利納斯,紐約大學醫學院神經學的研究人員之一,他這樣解釋道,「這一假設是保守的(在進化過程中),因為它是一種很智能的解決方案。由於系統涉及運動協調,因此我們希望一個機器擁有先進的電機控制,那麼(用來模擬的電路的)進行選擇將會很容易。」
他們的模擬器的一個獨特的方面是它使用模擬電路。類似於密德對大腦區域仿真的開創性研究,研究人員發現,通過在模擬模式中使用晶體管可以實現更少的組件而達到更高的性能。
其中一個小組的研究人員是美國西北大學的神經學家,費爾南多·穆薩-伊瓦爾迪,他評論了一個人造小腦電路在幫助殘疾人上的應用:「想想一個癱瘓的病人,可以設想很多尋常的事情,例如取一杯水、穿衣、脫衣,如果有一個機器人可以幫助他,就可以使病人更加獨立。」
高級別的理解功能:倣傚、預言和情感
思想的運作就像是戰鬥中的騎士——嚴格地控制著自己的數量,他們需要馬匹,並且只在關鍵的時刻派上用場。
——阿爾弗雷德·諾斯·懷特海德
人類智能的顯著特點不是運作的時候它在做什麼,而是停止的時候它在做什麼。
——馬文·明斯基
如果愛是一個答案的話,你能為這個答案尋找一個問題嗎?
——莉莉·湯姆林
因為它位於神經層次的頂端即大腦皮層,這是人類知之甚少的大腦部分。這個區域由六個位於大腦半球的最外層薄層組成,其中包含了數十億個神經元。根據索爾克研究所計算神經學實驗室的小托馬斯·巴托爾的生物學研究,「1立方毫米的大腦皮層可能包含了5億個有秩序的不同形狀和大小的神經突觸。」大腦皮層負責知覺、規劃、決策以及我們認為是意識性思維的大部分。
我們能夠使用語言——這個我們人類物種的另一個特有的屬性似乎就位於大腦皮層區域中。這裡有一個有趣的線索,是關於語言起源和促使區別性技能形成的關鍵性進化變革的:我們觀察到只有少數靈長類動物能夠使用(實際的)鏡子去掌握技能,如人類和猴子。理論家吉亞科莫·裡佐拉蒂和邁克爾·阿比布提出假設:語言起源於做手勢(猴子會,當然人類也會)。做手勢要求能夠在思想上將動作和對自己手勢的觀察相關聯。111他們的「鏡子系統假說」中語言進化的關鍵是一個叫做「等價」的性質,所謂「等價」是指對於做手勢的一方和接受手勢的一方對手勢(或者言論)的理解有著相同的意義。這就解釋了為什麼你在鏡子中所看到的和某個觀察你的人所看到的是一樣的(儘管要左右顛倒)。其他的動物不能夠以這種方式理解鏡子中的影像並且可以相信它們失去了等價這一關鍵能力。
一個與此緊密相關的概念是倣傚他人動作的能力(或者像人類嬰兒學發聲的例子)對語言的發展至關重要。112模仿要求能夠將看到的演示分解成幾個部分,每個部分都能夠通過遞歸和迭代的改進來掌握。
在語言能力學的新理論中,認為遞歸是一項關鍵能力。在諾姆·喬姆斯基的人類語言早期理論中,他引用了許多共同屬性來解釋人類語言的眾多相似之處。在2002年由馬克·豪澤、諾姆·喬姆斯基和特庫姆塞·費奇所撰寫的論文中,作者引用了遞歸這一單一屬性來證明人類物種獨特的語言天賦。113遞歸是將多個小的部分組合成一個大的部分,並且將這個大部分作為其他結構的一部分來使用,然後反覆繼續這個過程的能力。通過這種方式,我們就能夠以有限個單詞建立句子和段落的詳細結構。
另一個重要的人類大腦特徵是預測的能力,包括預測我們的決定和行為的結果。一些科學家認為預測是大腦皮層的主要功能,儘管小腦在預測活動中也起著重要作用。
有趣的是,我們能夠預測或者預期我們自己的決定。加利福尼亞大學戴維斯分校的生理學教授本傑明·裡貝特的工作顯示,發起一個行為的神經活動比大腦做出採取行動的決定要早1/3s。言外之意,裡貝特認為決定是真實存在的錯覺,即「意識跳出了循環」。認知科學家和哲學家丹尼爾·代尼特描述的現象如下:這一行為由大腦的某個部分加速,飛速把信號發送給肌肉,並且在途中暫停告訴你的意識代理將要發生什麼。114
最近進行的一項相關的實驗是,神經生理學家用電信號刺激大腦上的點位來產生特定的情感。這一課題隨即給出了經歷那些情感的解釋。眾所周知,那些左右大腦無法溝通的患者,大腦的其中一半(通常是左半腦)會對由另一半腦引起的動作給出複雜的解釋(「虛構症」),就像左半腦是右半腦的公共關係經紀人一樣。
人類大腦最複雜的能力是我們的情商,我將它視為大腦最尖端的技能。我們能夠恰當地感知回應情感,在社會環境中做出反應,有道德,聽得懂笑話並且能感知藝術音樂,這些都是大腦有內部連接的複雜的高級功能。很明顯,低級的感知與分析功能會參與到大腦的情感處理過程中,但是我們已經開始理解大腦的這個區域甚至能夠模擬特定的神經元去解決這些問題。
這些最近的解釋可以幫助我們去理解人類大腦與其他哺乳動物差異。答案很微小但卻很關鍵,這些能幫助我們理解大腦怎樣處理情緒和一些相關的情感。一個區別點是人類的大腦具有非常大的腦皮層,使我們擁有很強的規劃、決策和一些相關的分析思維能力。另外一個重要的區分點是情感被一種叫做梭形細胞(只在人類和類人猿中發現)的特殊細胞所控制,這些細胞非常大,而且有很長的神經細絲(稱作尖端突觸),這些神經細絲傳遞著來自很多其他大腦區域的特異性信號(見圖4-8)。隨著人類進化進程的深入,這種特殊神經元提供的廣泛連接或者這種深層次的連接變得越來越顯著。考慮到感情的複雜性,我們不會為梭形細胞建立深層連接以幫助我們處理情感和道德判斷而感到驚訝。
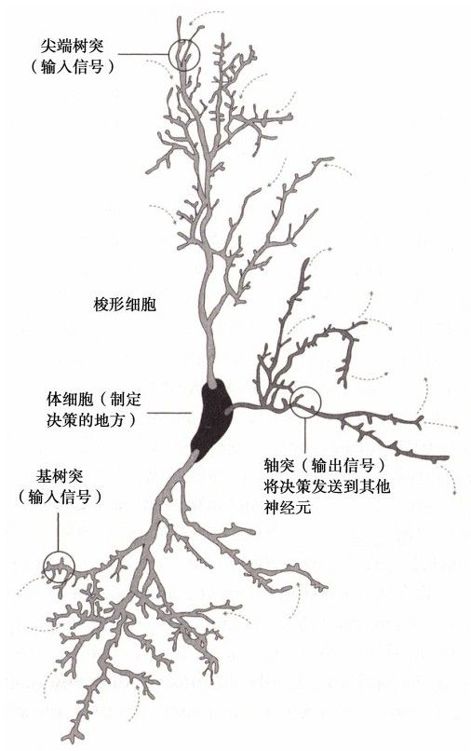
圖 4-8
令我們驚奇的是,在這狹小的區域裡只有極少量的梭形細胞,人類大腦中大概只有80000個(45000個在右半球,35000個在左半球),雖然差別很小,但是這些不同似乎能夠解釋為什麼情商是由右腦控制。大猩猩擁有約16000個梭形細胞,倭黑猩猩有2100個,黑猩猩有1800個。其他哺乳動物則完全沒有。
鳳凰城巴羅神經學研究所的亞瑟·克雷格博士最近提供了一個關於紡錘體細胞結構的描述。115來自皮膚、肌肉、組織和其他地方的神經輸入(估計在數百兆每秒)流進脊髓上部。這些神經攜帶的信息涉及觸覺的、溫度的、酸度的(例如,在肌肉中的乳酸)、食物通過胃腸道時的蠕動,以及許多其他類型的信息。這些數據會在腦幹和中腦中進行處理。稱作Lamina 1的關鍵神經元細胞繪製了一幅反映身體當前狀態的地圖,就像飛行控制者用來跟蹤飛機的顯示器。
然後信息會流過一個稱為後腹內側核(VMpo)的螺母大小的區域,它似乎能夠推斷出身體狀態的一些複雜反應,例如,「這個嘗起來很糟糕」,「好臭」,或者「光好刺眼」。這些日益複雜的信息最終在兩個稱為島狀皮層的區域結束。這些小指大小的結構位於皮層的左右兩側。克雷格把VMpo和兩個島狀區域描述為「代表物質我的一個系統」。
雖然機制尚未明確,但是這些區域對於自我意識和複雜情感是至關重要的。它們在其他動物身上更小。例如,VMpo在獼猴身上差不多只有一粒沙子大小,在低級動物中就更小。根據愛荷華大學的安東尼奧·達瑪西奧博士的觀點,這些發現越來越達成一個這樣的共識,那就是我們的情感和那些包含身體地圖的大腦區域是緊密聯繫的。116他們也與這樣的觀點達成一致,即我們很多的想法直接針對於身體,保護和提高自身,積極滿足它們無數的需求和慾望。
最近,我們已經發現從身體上以感知信息開始的另外一個層面上的過程。從兩個腦區來的數據傳輸到右腦前面的一個叫做前島葉的微小區域中。這是一個存在梭形細胞的區域,tMRI掃瞄為我們揭示出這個區域在人類處理像愛情、憤怒、憂傷和性慾的高層次的情感時會變得非常活躍。目標人物看著他的伴侶或者聽到自己孩子的哭聲時,梭形細胞也會受到刺激。
人類學家認為梭形細胞第一次出現是在1000~1500萬年前,在一種未被發現的常見的類人猿和猿人祖先中,他們的數量在10萬年前迅速增長。有趣的是,新生兒沒有梭形細胞,這些細胞在小孩4個月大的時候開始出現,並在1~3歲時迅速增長。幼兒對道德問題與複雜感情(比如愛)的處理和感知能力也是在這個時期發育的。
梭形細胞是通過它們長的頂端樹突與其他大腦部位的深入聯繫來獲得能量的。因此,頂端樹突所處理的高水平情感活動受感覺和認知領域的影響,對梭形細胞的具體活動方法進行逆向工程是很困難的,正因如此,只有掌握與它們相連接的其他區域的更好模型才能夠實現。然而,令我們意外的是,專門處理這種情緒的神經元非常少。小腦有500億神經元幫助我們處理技能的形成,大腦皮層中有數十億神經元用來轉換認識和理性規劃,但卻僅有八萬梭形細胞處理高級情感活動。必須指出的很重要的一點是,梭形細胞並不做合理分析,這就是為什麼我們在音樂與愛情面前沒有理性。然而,大腦的其他區域則忙於賦予我們神秘的高級情感活動以真正的意義。